Introducción
La pregunta por el origen de la vida es una de las más antiguas que ha formulado el pensamiento humano. Durante milenios, las culturas recurrieron a mitos de dioses creadores y relatos sagrados para explicar cómo lo inerte se convertía en viviente. Pero con Aristóteles comenzó un camino distinto: la búsqueda de una explicación natural, aunque equivocada, que sustituyera el mito por la observación. <sup>7</sup>
Ese trayecto nos conduce desde la generación espontánea medieval hasta la biogénesis de Pasteur, desde la intuición de Darwin sobre un “charco caliente” hasta la hipótesis de Oparin y Haldane, y desde los matraces de Miller y Urey hasta la reconstrucción de LUCA como ancestro común de todo lo viviente. Es un recorrido que revela la fuerza de la ciencia para transformar preguntas metafísicas en problemas experimentales.
Pero el tema no se agota en la biología. El origen de la vida conecta con la astrobiología contemporánea y con la paradoja de Fermi: la química parece empujar hacia la vida en cualquier rincón del cosmos, y sin embargo la conciencia reflexiva —la que pregunta por sí misma— parece ser rara, quizá única. Así, preguntarse por el inicio de la vida es también preguntarse por el lugar que ocupamos en un universo que puede estar lleno de microbios, pero vacío de voces.
Este ensayo explora esa historia: de Aristóteles a LUCA, de los virus como sombras del mundo de RNA al hallazgo de posibles biosignaturas en Marte, y de la universalidad de la biología a la fragilidad de la conciencia. Porque entender cómo empezó la vida no es solo un problema científico, sino una forma de comprender lo que somos y lo que, tal vez, nunca más vuelva a repetirse en el cosmos.
Aristóteles y la generación espontánea
Antes de Aristóteles, las culturas solían explicar la aparición de los seres vivos recurriendo a mitos de dioses creadores, fuerzas primordiales o cosmogonías fantásticas. Sin embargo, en el siglo IV a. C., Aristóteles se atrevió a formular una teoría naturalista que, aunque errónea, desplazaba el peso de la explicación desde el mito hacia la observación.

Su hipótesis más influyente fue la generación espontánea: la idea de que ciertos organismos podían surgir directamente de la materia inerte bajo determinadas condiciones. Así, gusanos y moscas podían brotar de la carne en descomposición; los ratones, de los graneros húmedos; y hasta ciertos peces, de la arena y el fango. Aristóteles pensaba que la materia contenía una suerte de principio vital latente, que se activaba por el calor y la humedad.
Aunque hoy sabemos que la biología refutó esta concepción, su importancia histórica es enorme. La generación espontánea representó el intento más sistemático de su tiempo por explicar el origen de la vida sin acudir a fuerzas sobrenaturales. En cierto modo, fue el primer eslabón en la larga cadena de teorías científicas que buscan responder la misma pregunta fundamental: ¿cómo surge la vida a partir de lo inerte?
Interludio: Parménides, el cambio y la generación espontánea
Parménides, en el siglo V a. C., defendía que el ser es uno, eterno e inmutable. Según él, el cambio es ilusorio: lo que existe no puede “nacer” de la nada ni “convertirse” en otra cosa, porque el no-ser no existe. Esta posición radical negaba la posibilidad de transformación real en la naturaleza.
Aristóteles, sin embargo, construyó toda su filosofía de la naturaleza contra esa negación. Para él, el cambio es real y necesario, pero requiere ser explicado con categorías lógicas: materia, forma, potencia y acto. Así, una semilla puede transformarse en árbol porque tiene en potencia la forma de árbol, y esa forma se actualiza mediante el calor, la nutrición y el tiempo.
La generación espontánea encajaba dentro de esta visión dinámica: aunque pareciera que la vida surgía “de la nada”, en realidad Aristóteles creía que la materia inerte guardaba en potencia la forma vital. El calor y la humedad no creaban el ser de la nada, sino que actualizaban una posibilidad ya inscrita en la materia.
En ese sentido, la teoría aristotélica puede verse como un puente entre Parménides y Heráclito: no niega el cambio (como Parménides), pero tampoco lo concibe como flujo absoluto (como Heráclito). Lo explica como un tránsito ordenado de la potencia al acto.
La Edad Media y el Renacimiento: la persistencia de la generación espontánea
Durante la Edad Media, la teoría aristotélica de la generación espontánea se integró en el gran edificio del pensamiento escolástico. Los filósofos y teólogos cristianos, como Santo Tomás de Aquino, aceptaban que Dios había creado el mundo, pero también admitían que dentro de ese mundo existían procesos naturales autónomos. En ese sentido, la generación espontánea no se veía como una negación de lo divino, sino como un mecanismo que Dios mismo había dispuesto para mantener la vida. La humedad, el calor y la putrefacción eran considerados “instrumentos” de la creación continua.

En la práctica, los manuales médicos y de historia natural medievales recogían observaciones que parecían confirmar la idea: insectos saliendo de cadáveres, gusanos apareciendo en frutas, ranas “brotando” en el lodo tras la lluvia. La experiencia cotidiana reforzaba el dogma aristotélico, y nadie veía necesidad de cuestionarlo.
Con el Renacimiento, lejos de desaparecer, la creencia en la generación espontánea se revitalizó. El redescubrimiento de Aristóteles y la pasión por la experimentación le dieron nuevos bríos. Autores como Paracelso llegaron a proponer recetas “alquímicas” para generar vida: desde fabricar ratones a partir de granos de trigo y ropa sucia, hasta la creación artificial de un homúnculo dentro de un frasco de vidrio. Aunque estas ideas hoy parecen fantasiosas, en su momento formaban parte del espíritu renacentista: la convicción de que la naturaleza podía ser manipulada, reproducida y comprendida a través del ingenio humano.
Así, desde los monasterios medievales hasta los laboratorios alquímicos del Renacimiento, la generación espontánea se mantuvo como explicación oficial durante casi dos milenios. Fue necesario el choque de la nueva ciencia experimental del siglo XVII para empezar a resquebrajarla.
Louis Pasteur y la caída de la generación espontánea
En el siglo XIX, tras siglos de aceptación, la teoría de la generación espontánea todavía tenía defensores. La observación cotidiana parecía confirmarla: caldo de carne que se llenaba de bacterias, infusiones que fermentaban por sí solas, mohos que aparecían en el pan. Para muchos naturalistas, la vida seguía brotando espontáneamente allí donde hubiera humedad y nutrientes.
Fue Louis Pasteur quien, con una serie de experimentos rigurosos entre 1859 y 1864, desmontó de raíz esa creencia. Su estrategia fue simple pero genial: demostrar que los microorganismos que aparecían en los caldos no surgían de la materia inerte, sino que procedían del ambiente.
En su célebre experimento de los matraces de cuello de cisne, Pasteur hervía caldos nutritivos dentro de recipientes de vidrio cuyo cuello alargado y curvado permitía el paso del aire, pero atrapaba el polvo y las partículas. Mientras el cuello permaneciera intacto, el caldo se mantenía estéril indefinidamente; en cuanto se rompía y el polvo del aire entraba, la vida microbiana brotaba de inmediato. La conclusión era inequívoca: la vida no se genera de la nada, sino de otra vida preexistente (omne vivum ex vivo).
Con esto, Pasteur no solo derribó una creencia milenaria, sino que estableció las bases de la biogénesis moderna y de la microbiología experimental. Su triunfo científico coincidió además con el auge del darwinismo, lo que abrió una pregunta mucho más profunda: si la vida siempre proviene de otra vida, ¿cómo surgió la primera célula?
El “pequeño charco caliente” de Darwin
Louis Pasteur había demostrado experimentalmente que la vida, en las condiciones actuales de la Tierra, no surge de la materia inerte. Pero Darwin no podía ignorar la pregunta de fondo: si la vida siempre procede de otra vida, ¿cómo apareció la primera?
n una carta privada a su amigo Joseph Hooker (1871), Darwin se permitió especular. Imaginó que, en los inicios del planeta, en “un pequeño charco caliente” (a warm little pond), con presencia de amoníaco, sales minerales, luz solar, calor eléctrico y otras formas de energía, podrían haberse formado compuestos químicos cada vez más complejos. Si esos compuestos adquirían la capacidad de replicarse y evolucionar, entonces el problema del origen quedaba resuelto en términos naturales.

Darwin nunca publicó esta hipótesis en sus libros, quizá porque temía que fuera usada por sus críticos como un flanco débil. Sin embargo, dejó planteada la posibilidad de una abiogénesis natural en el pasado remoto, distinta de la generación espontánea cotidiana que había refutado Pasteur. Era un marco conceptual nuevo:
- No generación espontánea aquí y ahora, sino un origen de la vida en condiciones muy específicas y excepcionales de la Tierra primitiva.
- Una primera chispa química que, una vez encendida, ya no necesitó repetirse porque la evolución darwiniana tomó el relevo.
Este “pequeño charco caliente” es considerado hoy el germen de la hipótesis prebiótica, y sería retomado en el siglo XX por Alexander Oparin en Rusia y John Haldane en Inglaterra, quienes dieron forma a la teoría moderna del origen químico de la vida.
Alexander Oparin y la “Sopa Prebiótica”
La intuición de Darwin sobre el origen de la vida encontró un eco inesperado décadas más tarde en Rusia. Allí, el botánico Kliment Timiriazev, discípulo y difusor entusiasta de las ideas de Darwin, se convirtió en el gran puente entre el darwinismo británico y la ciencia rusa. Uno de sus alumnos más brillantes, Alexander Oparin, heredó esa visión y la llevó a un nuevo nivel: inspirado por el marco evolutivo darwiniano y guiado por la enseñanza de Timiriazev, elaboró en los años veinte la hipótesis de la evolución química. Así, una simple especulación epistolar se transformó, a través de un linaje intelectual, en la primera teoría científica sistemática sobre el origen de la vida.

Haldane y la abiogénesis
Mientras en la Rusia de los años veinte Oparin proponía que la vida surgió a partir de una “sopa prebiótica” de compuestos orgánicos acumulados en la Tierra primitiva, en Inglaterra, de manera independiente, el genetista y bioquímico J.B.S. Haldane (1892–1964) llegó a conclusiones muy similares.
En un ensayo de 1929 titulado The Origin of Life, Haldane especuló que la atmósfera primitiva de la Tierra —carente de oxígeno libre y rica en gases reductores como metano, amoníaco e hidrógeno— favoreció la síntesis de moléculas orgánicas bajo la acción de la radiación solar ultravioleta. En esa “caldera química”, los océanos actuaron como un gigantesco laboratorio natural en el que se formaron azúcares, aminoácidos y bases nitrogenadas, precursores de la vida.

Para Haldane, el paso clave fue la aparición de sistemas coloidales, a los que llamó coacervados, que podían concentrar moléculas y facilitar reacciones químicas. Aunque no eran organismos vivos, representaban un estadio intermedio entre la química inorgánica y la biología.
Lo notable es que, sin conocerse entre sí, Oparin en Moscú y Haldane en Londres convergieron en una misma intuición: la vida no había surgido por un acto milagroso ni por generación espontánea contemporánea, sino mediante un proceso lento y natural de evolución química en la Tierra primitiva. Esa coincidencia marcó el inicio de la teoría moderna del origen de la vida.
El experimento de Miller y Urey (1953): poner a prueba a Oparin y Haldane
Las ideas de Oparin y Haldane sobre la evolución química permanecieron durante décadas en el terreno de la especulación teórica. No existían pruebas experimentales que demostraran que, a partir de condiciones similares a las de la Tierra primitiva, pudieran surgir compuestos orgánicos básicos para la vida. Esa prueba llegó en 1953, gracias al joven químico Stanley Miller, bajo la supervisión de Harold Urey en la Universidad de Chicago.
Miller diseñó un aparato cerrado que simulaba la atmósfera primitiva según el modelo de Haldane: una mezcla de metano, hidrógeno, amoníaco y vapor de agua, sin oxígeno libre. Hizo circular descargas eléctricas continuas, imitando la energía de los rayos en la atmósfera primitiva, y mantuvo la mezcla en constante circulación sobre agua caliente que simulaba los océanos.
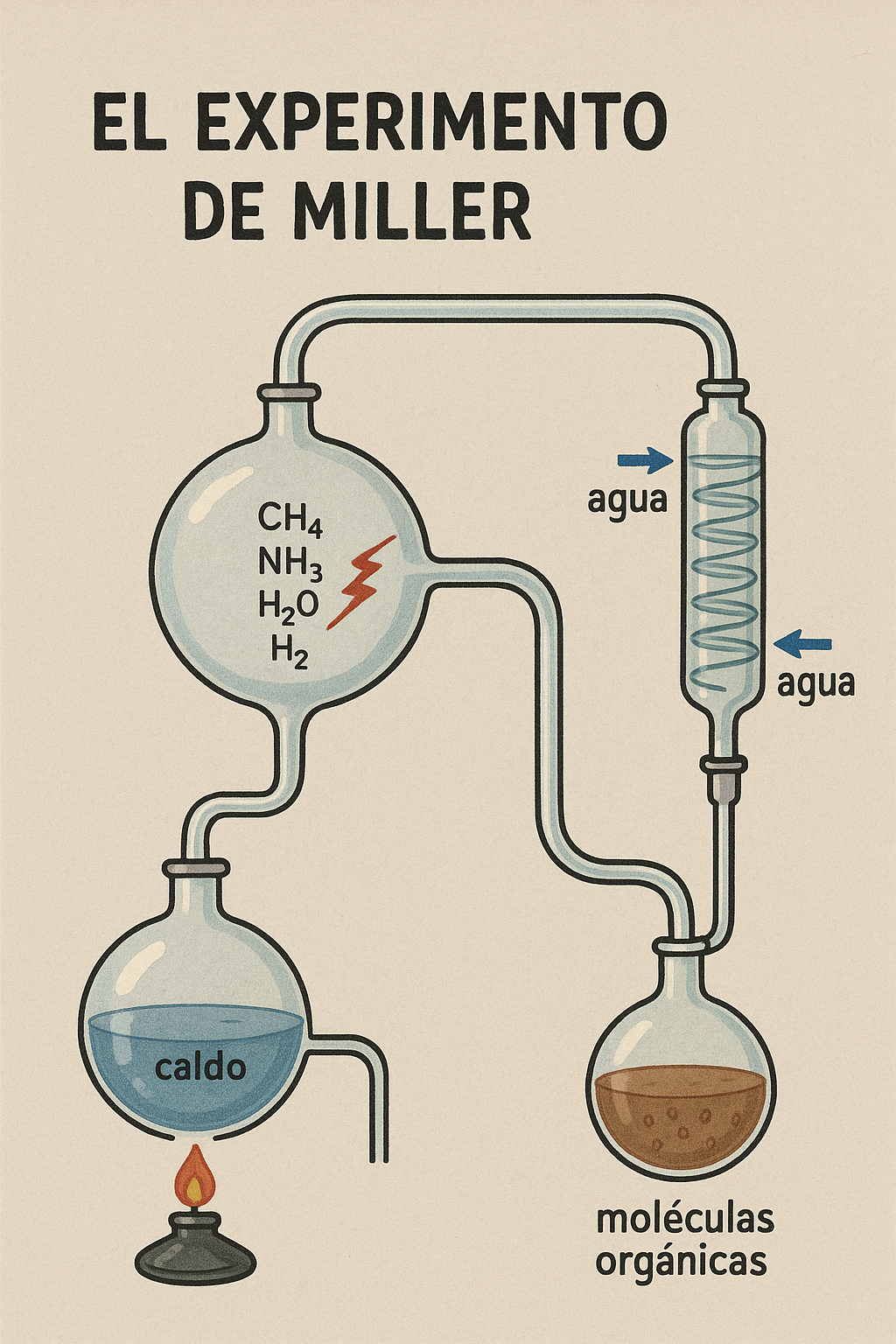
Al cabo de una semana, el líquido transparente se había vuelto marrón rojizo. Los análisis revelaron algo sorprendente: en el “caldo” se habían formado aminoácidos —los bloques fundamentales de las proteínas— junto con otros compuestos orgánicos esenciales. Era la primera confirmación experimental de que la materia inorgánica podía transformarse en materia orgánica compleja bajo condiciones plausibles de la Tierra primitiva.
El experimento de Miller–Urey no creó vida, pero demostró que el camino propuesto por Oparin y Haldane era científicamente viable. Desde entonces, se abrió un campo entero de investigación sobre la química prebiótica, que hoy abarca desde simulaciones en laboratorio hasta estudios sobre meteoritos y entornos planetarios.
Meteoritos condritos carbonáceos: cápsulas del origen de la vida
La explicación moderna del origen de la vida no se limita a la Tierra como escenario aislado. Los estudios de meteoritos condritos carbonáceos —fragmentos primitivos del sistema solar— han revelado que estos cuerpos celestes trajeron consigo tanto agua como moléculas orgánicas complejas, indispensables para el arranque de la biología.

La diferencia fundamental radica en su historia geológica. Mientras la Tierra primitiva sufrió un proceso de diferenciación interna —altas presiones y temperaturas que fundieron su interior, separando el núcleo metálico del manto y la corteza—, muchos de estos meteoritos no llegaron a fundirse nunca. Son materiales “no procesados”, que conservaron intacta la química del disco protoplanetario de hace 4.600 millones de años.

Gracias a ello, los condritos preservan moléculas que en la Tierra habrían sido destruidas: aminoácidos, hidrocarburos, ácidos carboxílicos e incluso compuestos que recuerdan a bases nitrogenadas. Ejemplos famosos, como el meteorito Murchison caído en Australia en 1969, muestran que el espacio interplanetario ya contenía la “materia prima” para la vida.
Este hecho cambia el marco de la pregunta: los ladrillos básicos de la biología no tuvieron que sintetizarse todos en la Tierra; algunos llegaron desde fuera, transportados por meteoritos y cometas. En ese sentido, la abiogénesis terrestre se alimentó de un aporte cósmico.
La tierra hace 4500 millones de años
Hace unos 4.500 millones de años, la joven Tierra —todavía en proceso de formación dentro del disco protoplanetario— sufrió un choque colosal con un protoplaneta al que los astrónomos llaman Teia. El impacto liberó una cantidad de energía equivalente a miles de millones de bombas atómicas, fundiendo grandes porciones de la corteza y el manto terrestre. Este hecho tuvo como consecuencias:
- Formación de la Luna. A consecuencia del impacto parte del material expulsado al espacio se acumuló en órbita y, con el tiempo, dio lugar a nuestro satélite natural.
- Oceano de magma: la Tierra quedó cubierta por un océano global de magma, con temperaturas extremas, atmósfera casi inexistente y una superficie constantemente bombardeada por asteroides y cometas.
- Atmósfera primitiva: la colisión barrió cualquier atmósfera previa. La nueva atmósfera, producto de la desgasificación volcánica, estaba compuesta por dióxido de carbono, vapor de agua, monóxido de carbono, nitrógeno, amoníaco y metano, pero sin oxígeno libre.
- Ciclo del agua: al enfriarse lentamente, el vapor de agua condensó y dio origen a los primeros océanos líquidos, mientras que los impactos de cometas y meteoritos aportaron agua adicional y compuestos orgánicos.
En este escenario extremo, la superficie terrestre se convirtió en un laboratorio gigantesco. La ausencia de oxígeno libre evitaba la destrucción inmediata de moléculas frágiles, y las abundantes fuentes de energía —rayos, radiación ultravioleta, calor volcánico, impactos meteóricos— impulsaban reacciones químicas que, poco a poco, generaban los bloques orgánicos de la vida.
Así, tras la violencia del impacto de Teia, la Tierra entró en una etapa crítica: la de la evolución química prebiótica, donde moléculas simples comenzaron a ensamblarse en formas más complejas, abriendo el camino hacia el RNA. Sin embargo, hubo de atravesar otra etapa crítica El Bombardeo Intenso Tardío (LHB).
Un planeta hostil pero fértil: El Bombardeo Intenso Tardío
El bombardeo intenso tardío fue un período clave en el que los asteroides condríticos carbonáceos, y en menor medida los cometas, aportaron agua, compuestos orgánicos y minerales esenciales a la Tierra primitiva. Estos materiales sentaron las bases para la evolución química, proporcionando los ingredientes necesarios para formar moléculas complejas y, eventualmente, los primeros sistemas biológicos. Aunque no fue el único evento que contribuyó, su impacto fue significativo para crear las condiciones que hicieron posible el origen de la vida.

Se cree que fue causado por inestabilidades en la órbita de los planetas gigantes (Júpiter y Saturno), que desestabilizaron millones de asteroides y cometas, enviándolos hacia el interior del sistema solar.
Durante unos 300 millones de años, la Tierra y la Luna fueron golpeadas por una lluvia constante de meteoritos de todos los tamaños. Las cicatrices aún se observan en la superficie lunar, ya que allí no hubo erosión ni tectónica que borrara las huellas.Cada impacto liberaba enormes cantidades de energía, fundiendo regiones de la corteza y evaporando océanos temporales.
Sin embargo, al mismo tiempo, estos cuerpos aportaban agua adicional y moléculas orgánicas complejas (aminoácidos, bases nitrogenadas, hidrocarburos), enriqueciendo la química de la superficie terrestre.Se piensa que hubo ciclos de destrucción y regeneración: los impactos esterilizaban grandes zonas, pero también sembraban materia prima y generaban nichos hidrotermales donde la química prebiótica podía avanzar.
Paradójicamente, aunque el Bombardeo Intenso Tardío hacía de la Tierra un planeta hostil para cualquier forma de vida establecida, también la convirtió en un laboratorio fértil para la evolución química. La combinación de agua, compuestos orgánicos extraterrestres y fuentes abundantes de energía (impactos, volcanes, radiación) creó un caldo propicio para que, más adelante, se gestara el mundo de RNA.
¿Cuándo apareció la vida?
Estudios recientes publicados en Nature Ecology & Evolution en 2024 han modificado radicalmente el origen de la vida. Según estos análisis moleculares, el Último Ancestro Común Universal (LUCA) ya existía hace unos 4.2 mil millones de años, es decir, antes del Bombardeo Intenso Tardío. Esta conclusión tiene implicaciones profundas: significa que la evolución química, ese proceso que condujo desde moléculas simples hasta la formación del RNA —el verdadero culmen de la química prebiótica—, tuvo lugar en una Tierra todavía joven, apenas estabilizada tras el impacto con Teia.
La consecuencia es reveladora: la vida no esperó a que pasara la tormenta cósmica; surgió en medio de ella. El Bombardeo Intenso Tardío, lejos de ser el origen de la vida, habría representado más bien su primera gran amenaza de extinción. Las formas prebióticas o protocelulares que ya existían pudieron verse diezmadas o incluso casi borradas, y sin embargo, sobrevivieron lo suficiente como para dejar un linaje: aquel que reconocemos hoy en LUCA..
Así, el descubrimiento actual obliga a adelantar la fecha de la formación del RNA —el esqueleto informacional de la vida— a un momento anterior a los 4.2 mil millones de años, en una Tierra que aún llevaba las cicatrices ardientes del impacto de Teia y que pronto sería azotada por una lluvia interminable de rocas cósmicas.
Cronología temprana del origen de la vida
- Formación de la Tierra (hace 4.5 mil millones de años): nuestro planeta se originó a partir de la acreción de polvo y rocas del disco protoplanetario que rodeaba al Sol joven. Poco después sufrió el impacto de Teia, que dio origen a la Luna y dejó a la Tierra cubierta por un océano de magma.
- Enfriamiento y estabilización (~4.2 Ga): tras unos 300 millones de años, la superficie comenzó a solidificarse, el vapor de agua condensó formando océanos y la atmósfera primitiva se estableció, rica en CO₂, metano, amoníaco y nitrógeno, pero sin oxígeno libre.
- LUCA temprano (~4.2 Ga): de acuerdo con estimaciones moleculares recientes (Nature Ecology & Evolution, 2024), pudo existir ya un Último Ancestro Universal Común (LUCA), lo que sugiere que la vida apareció muy pronto, apenas el planeta ofreció estabilidad suficiente para sostenerla.
- Bombardeo Intenso Tardío (4.1–3.8 Ga): una lluvia de meteoritos y cometas golpeó la Tierra durante cientos de millones de años. Este episodio pudo haber causado la primera extinción masiva prebiótica, eliminando formas tempranas de vida o reduciéndolas a refugios muy localizados. Al mismo tiempo, aportó agua y moléculas orgánicas desde el espacio, enriqueciendo los océanos.
- Resurgimiento de la vida (~3.8 Ga): una vez que el bombardeo remitió, los océanos y la atmósfera ofrecieron condiciones más estables. En este contexto se consolidó la evolución química que llevó a las primeras formas de RNA y a los ancestros de todos los organismos actuales.
¿Cuáles fueron las primeras formas de vida?
Durante gran parte del siglo XX, la hipótesis de Oparin y Haldane dominó las explicaciones sobre el origen de la vida. Según ellos, la secuencia lógica era imaginar primero la formación de estructuras semejantes a células —pequeños glóbulos coloidales o coacervados rodeados por membranas rudimentarias— que luego habrían incorporado progresivamente la capacidad de almacenar y transmitir información genética mediante moléculas como el DNA. Bajo esta visión, la compartimentación antecedía a la información hereditaria.
Sin embargo, a partir de la década de 1980 surgió una hipótesis revolucionaria: la del mundo de RNA. Experimentos y descubrimientos bioquímicos revelaron que el RNA no es solo un intermediario pasivo entre el DNA y las proteínas, sino una molécula capaz de almacenar información y, al mismo tiempo, catalizar reacciones químicas. En otras palabras, el RNA podía actuar como replicador y como enzima (ribozima). Esto abría un nuevo escenario: antes de que existiera vida (células con DNA estable y metabolismo), moléculas de RNA autorreplicantes pudieron ser las primeras entidades “vivas” en sentido funcional.
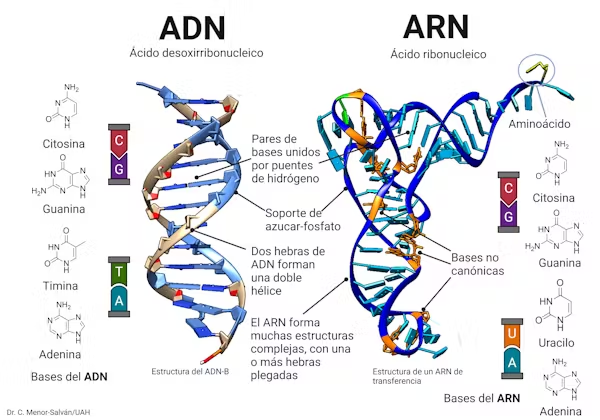
El cambio de paradigma es profundo. En vez de imaginar primero vesículas y membranas protectoras, la hipótesis del mundo de RNA propone que la chispa de la vida comenzó con replicadores moleculares capaces de copiarse a sí mismos, aunque de forma imperfecta. La membrana celular habría llegado después, como una innovación evolutiva que otorgó mayor estabilidad y aislamiento a esas moléculas replicantes.
En esta visión moderna, la vida no surgió como una célula completa desde el inicio, sino como un proceso: primero las moléculas replicantes (RNA), luego la membrana protectora, y mucho más tarde el sofisticado sistema DNA–proteínas que hoy caracteriza a todos los organismos.
Evidencias que sustentan el mundo de RNA
En conjunto, estas evidencias refuerzan la hipótesis de que antes de existir DNA estable y proteínas eficientes, hubo un mundo de RNA donde las primeras moléculas replicantes se copiaban, variaban y evolucionaban. Solo más tarde, la evolución incorporó la membrana celular y, finalmente, el sistema DNA-proteínas, mucho más robusto.
- Ribozimas:
El descubrimiento de que algunas moléculas de RNA tienen capacidad catalítica (es decir, actúan como enzimas) fue un parteaguas. Estas ribozimas pueden cortar, unir y copiar fragmentos de RNA. Esto demostró que el RNA podía cumplir la doble función necesaria para la vida primitiva: información + catálisis. - Síntesis prebiótica de nucleótidos:
Experimentos recientes han mostrado que, bajo condiciones plausibles de la Tierra primitiva (radiación ultravioleta, minerales como arcillas, ciclos de hidratación y deshidratación), es posible formar nucleótidos —los ladrillos básicos del RNA— a partir de moléculas simples. Aunque todavía no tenemos un camino completo, se ha avanzado en demostrar que las piezas podían surgir de manera natural. - Autorreplicación parcial:
Se han diseñado sistemas en laboratorio donde cadenas cortas de RNA pueden replicar parcialmente su propia secuencia con la ayuda de ribozimas. No es todavía un proceso perfecto ni completamente autónomo, pero apunta a que moléculas primitivas de RNA pudieron alcanzar cierto grado de autocopia rudimentaria en la Tierra temprana. - Conservación en la biología moderna:
El RNA sigue siendo fundamental en todas las células actuales:- Transporta información (ARNm).
- Actúa en la síntesis de proteínas (ARNt y ARNr).
- Regula procesos celulares (microRNAs).
- Forma el núcleo activo del ribosoma, la máquina traductora universal.
Esta centralidad sugiere que en algún momento remoto la vida dependía casi exclusivamente del RNA.
Escenarios geológicos para el origen del RNA
Es probable que no existiera un único “escenario” privilegiado, sino que varios ambientes de la Tierra primitiva —respiraderos profundos, charcas volcánicas y superficies minerales— funcionaran como laboratorios naturales en paralelo. En todos ellos, la química prebiótica pudo experimentar múltiples vías, hasta que alguna dio lugar a moléculas de RNA capaces de replicarse y abrir el camino hacia la vida.
- Respiraderos hidrotermales en el fondo oceánico:
- Estos sistemas, presentes aún hoy en las dorsales oceánicas, expulsan agua rica en minerales y gases reductores a temperaturas elevadas.
- Ofrecen gradientes de energía (química y térmica) que pudieron impulsar la síntesis de moléculas orgánicas.
- Las paredes porosas de los respiraderos, compuestas de minerales como sulfuros de hierro, actuaban como microcompartimentos naturales, concentrando compuestos y favoreciendo reacciones químicas.
- Algunos investigadores ven aquí el escenario más plausible para la transición de moléculas simples a replicadores de RNA.
- Charcas volcánicas poco profundas:
- En la superficie terrestre primitiva existían lagunas cercanas a volcanes, con ciclos de hidratación y desecación impulsados por el clima y la geotermia.
- Estos ciclos son fundamentales, porque la desecación concentra los solutos y permite que las moléculas se unan en cadenas más largas. Luego, al rehidratarse, esas cadenas pueden dispersarse y replicarse.
- Este ambiente también recibía abundante radiación ultravioleta, que aunque destructiva, podía haber jugado un papel catalítico en la formación de nucleótidos y enlaces.
- Superficies minerales y arcillas:
- Minerales como la montmorillonita han demostrado en laboratorio la capacidad de catalizar la polimerización de nucleótidos, es decir, unirlos en cadenas de RNA.
- Las superficies minerales también pudieron servir como “andamios” para estabilizar moléculas frágiles y mantenerlas en contacto.
- En este escenario, la geología misma funciona como catalizador, ayudando a que moléculas dispersas se organicen en polímeros replicantes.

De RNA a DNA: la gran transición
El mundo de RNA ofrecía una solución elegante al problema del origen: moléculas que podían almacenar información y, al mismo tiempo, catalizar reacciones químicas. Sin embargo, tenía limitaciones serias:
- El RNA es químicamente inestable, se degrada fácilmente en presencia de calor, radiación o incluso agua.
- Su capacidad catalítica es real, pero mucho más limitada que la de las proteínas, que ofrecen una enorme diversidad de formas y reactividades.
- La fidelidad en la replicación era baja, lo que generaba errores acumulativos.
Frente a esto, la evolución encontró una salida: la transición hacia un sistema más robusto y especializado. Este sistema, que parece natural hoy, fue en realidad el producto de una larga transición evolutiva desde un mundo en que el RNA hacía casi todo.
El DNA es químicamente más estable que el RNA, gracias a su doble hélice y a la ausencia del grupo hidroxilo en la posición 2’. Esto permitió almacenar genomas más grandes y complejos, con menor riesgo de degradación. Probablemente surgió mediante enzimas primitivas (ribozimas o proteínas tempranas) que comenzaron a copiar RNA en DNA, un proceso análogo a la transcriptasa inversa que todavía usan ciertos virus.
Una vez que aparecieron los primeros polipéptidos, la selección favoreció a los catalizadores más eficientes. Las proteínas, con sus veinte aminoácidos distintos, ofrecen una plasticidad estructural mucho mayor que el RNA. Poco a poco, las proteínas desplazaron al RNA de la mayoría de las funciones catalíticas. El resultado final fue una división del trabajo:
DNA como depósito estable de la información genética.
RNA como intermediario y regulador (ARNm, ARNt, ARNr).
Proteínas como la maquinaria catalítica y estructural dominante.
El RNA nunca desapareció del todo. El ribosoma —la máquina universal de traducción— sigue teniendo un núcleo de RNA catalítico, recordándonos que alguna vez la vida dependió exclusivamente de él. En cierto sentido, cada célula moderna lleva dentro un fósil viviente molecular de aquel mundo remoto.
Interludio 1: ¿Habia Selección Natural en el mundo de RNA?
Si imaginamos aquel mundo remoto dominado por moléculas de RNA, no podemos pensar en un escenario idílico de cooperación. La vida, desde sus orígenes más primitivos, estuvo marcada por la competencia y el parasitismo. Junto a los progenotes, esas entidades rudimentarias capaces de replicarse y mantener un conjunto básico de “protogenes”, surgieron secuencias de RNA más cortas, incompletas, incapaces de replicarse solas.
Estas secuencias, lejos de desaparecer, aprendieron a aprovechar la maquinaria de los progenotes, robando fragmentos de información genética y utilizándolos para perpetuarse. Eran, en cierto modo, los primeros virus: parásitos moleculares que vivían a expensas de otros replicadores.
Aunque pueda sonar negativo, este parasitismo temprano desempeñó un papel creativo. Al forzar a los progenotes a defenderse, aceleró la innovación evolutiva: obligó a desarrollar estrategias de copia más precisas, mecanismos rudimentarios de compartimentación y, quizás, dio un impulso a la aparición de membranas protectoras. En otras palabras, el “robo” de protogenes por parte de los virus primitivos pudo ser un motor oculto de la complejidad.
Hoy, los virus siguen siendo dependientes, pero al mismo tiempo se han convertido en agentes fundamentales de la evolución, portadores de genes entre especies y arquitectos invisibles de la diversidad. En ellos late la memoria de aquel mundo de RNA: sombras ancestrales que nunca dejaron de acompañar a la vida.
Interludio 2: El gen egoísta en el mundo de RNA
Cuando Richard Dawkins formuló la hipótesis del “gen egoísta” en los años setenta, no hablaba de egoísmo en un sentido moral, sino como metáfora de la lógica profunda de la evolución: lo que realmente compite en la vida no son los organismos completos, sino los genes, fragmentos de información cuya “meta” es perpetuarse. Los organismos no serían más que vehículos de esa misión.
Si llevamos esa idea al mundo de RNA, el panorama cobra una fuerza especial. Allí no existían todavía células complejas ni genomas organizados, sino “progenotes” cadenas de RNA que habían alcanzado la capacidad de autorreplicarse. No se trataba de un “individuo” buscando sobrevivir, sino de un conjunto de genes incipientes luchando por mantener la función que habían logrado.
Podemos imaginar, por ejemplo, un fragmento de RNA que codificaba para una ribozima capaz de reparar daños en la cadena. Ese “gen de reparación” no pensaba ni tenía intención, pero al contribuir a la estabilidad del conjunto aumentaba las probabilidades de que esa misma función se copiara y se transmitiera. De este modo, los genes útiles para la replicación y la supervivencia molecular tendían a conservarse, mientras que los que no aportaban nada se desvanecían en la sopa prebiótica.
Bajo esta luz, el mundo de RNA no fue un escenario caótico de moléculas azarosas, sino una primera expresión de la lógica del gen egoísta: secuencias que competían entre sí por persistir, que cooperaban cuando era ventajoso y que dieron origen, paso a paso, a las arquitecturas más complejas de la vida.
El Último Ancestro Común Universal (LUCA)
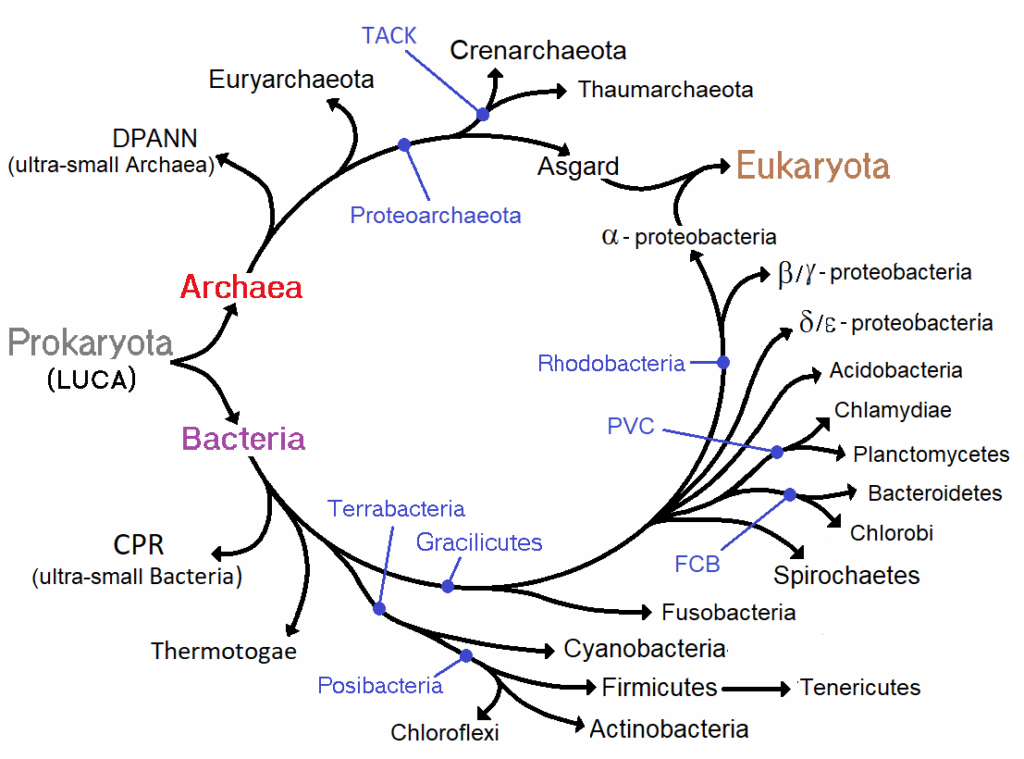
En la cultura popular suele hablarse del “eslabón perdido” como si la evolución fuese una cadena lineal donde falta una pieza intermedia. Esa idea es engañosa: la evolución no es una cadena, sino un árbol ramificado, donde cada bifurcación representa un ancestro común a partir del cual se diversifican distintas ramas.
En este marco, la cladística —la ciencia que clasifica a los organismos según relaciones de parentesco evolutivo— utiliza el concepto de Último Ancestro Común (UAC o LUCA, en el caso universal). El UAC no es un individuo fósil que podamos descubrir intacto, ni un “eslabón” que una mecánicamente dos especies. Más bien, es una reconstrucción teórica: el punto del árbol filogenético en el que convergen las ramas de dos o más linajes actuales.
Por ejemplo:
- El último ancestro común entre humanos y chimpancés no fue un “eslabón perdido” entre mono y hombre, sino una población real de primates de la que descendieron ambos linajes.
- De igual manera, el LUCA (Last Universal Common Ancestor) no fue la primera forma de vida, sino el punto más antiguo al que podemos retroceder siguiendo el linaje de todos los organismos vivos actuales.
Esto significa que:
- Nunca encontraremos al “eslabón perdido” porque la evolución no tiene eslabones sueltos, sino ramas divergentes.
- Los últimos ancestros comunes son conceptos dinámicos, reconstruidos a partir de comparaciones genéticas y cladísticas.
- En el caso de LUCA, hablamos de una población celular primitiva, no de un solo individuo. Esa población compartía un conjunto de genes y características que luego heredaron todas las formas de vida que existen hoy.
En síntesis, el concepto de Último Ancestro Común nos libera de la visión lineal de la evolución y nos obliga a pensar en términos de árboles, bifurcaciones y linajes, donde lo que importa no es el “eslabón perdido”, sino la reconstrucción del nodo ancestral que dio origen a la diversidad actual.
Reconstrucción de LUCA
Los estudios de genómica comparada han permitido inferir las características de LUCA al identificar los genes y proteínas que son comunes a todas las formas de vida actuales. No se trata de un fósil encontrado, sino de una reconstrucción teórica basada en la información genética que comparten bacterias, arqueas y eucariotas.
1. ¿Cuándo vivió LUCA?
- Los datos más recientes (Moody et al., Nature Ecology & Evolution, 2024) sugieren que LUCA existió hace unos 4.2 mil millones de años, es decir, antes del Bombardeo Intenso Tardío.
- Esto implica que la evolución química y el mundo de RNA se consolidaron muy pronto tras el enfriamiento de la Tierra.
2. ¿Qué tipo de organismo era?
- LUCA no fue el primer ser vivo, sino el último punto de convergencia entre todos los linajes actuales.
- Se trataba de una célula primitiva, probablemente sin la complejidad de las bacterias modernas, pero ya organizada.
- Los análisis genómicos sugieren que era un organismo anaerobio (vivía sin oxígeno, porque la atmósfera carecía de él) y quimiolitótrofo, es decir, que obtenía energía a partir de reacciones químicas con compuestos inorgánicos.
3. ¿Cómo era su metabolismo?
- Dependía del hidrógeno molecular (H₂) como fuente de electrones y del CO₂ como fuente de carbono.
- Su metabolismo se parece al de algunos microbios actuales que viven en respiraderos hidrotermales submarinos, lo que refuerza la idea de que esos ambientes pudieron ser su cuna.
- Poseía enzimas relacionadas con la síntesis de nucleótidos, aminoácidos y lípidos básicos, lo que indica que ya podía construir muchos de los bloques fundamentales de la vida.
4. ¿Qué sabemos de su genética?
- LUCA probablemente tenía un genoma basado en DNA, heredado del mundo de RNA pero ya más estable.
- Usaba RNA como intermediario y ribozimas en procesos clave.
- Poseía al menos unas 2,600 proteínas (según reconstrucciones modernas), lo que lo convierte en un organismo sorprendentemente sofisticado para su época.
- El código genético que usaba para traducir nucleótidos en aminoácidos era esencialmente el mismo que conservamos hoy, lo que muestra la enorme estabilidad evolutiva de este sistema.
5. ¿Dónde vivió LUCA?
La mayoría de hipótesis lo sitúan en ambientes ricos en minerales y gradientes químicos, probablemente:
- Respiraderos hidrotermales alcalinos en el fondo del océano, donde las reacciones entre agua y rocas ultramáficas generaban abundante hidrógeno.
- Nichos protegidos de la radiación cósmica y de los impactos, donde las células primitivas podían sobrevivir y diversificarse.
La diversificación de LUCA
LUCA no fue el primer ser vivo, pero sí el último ancestro común de todos los organismos actuales. Desde él se produjo una de las bifurcaciones más profundas de la historia de la vida: la separación en bacterias y arqueas, dos dominios celulares radicalmente distintos, de los cuales más tarde emergerían los eucariotas.
1. Bacterias: la rama pragmática
- Características: células pequeñas, con una gran diversidad metabólica. Incluyen organismos fotosintéticos (como las cianobacterias), fermentadores, reductores y oxidadores de múltiples compuestos.
- Innovación clave: la pared celular de peptidoglicano, que les dio resistencia y versatilidad.
- Impacto: fueron las primeras en colonizar todos los ambientes de la Tierra y, con la fotosíntesis oxigénica, transformaron la atmósfera, preparando el camino para la vida aeróbica.
2. Arqueas: la rama extrema
- Características: organismos unicelulares que, aunque se parecen a las bacterias en forma, son genéticamente muy distintos.
- Ambientes: muchas arqueas prosperan en condiciones extremas (altas temperaturas, alta salinidad, acidez), pero también habitan océanos, suelos y el intestino humano.
- Innovación clave: membranas celulares únicas, con lípidos distintos a los de bacterias y eucariotas, lo que les confiere gran resistencia.
- Impacto: mantienen un linaje paralelo y misterioso, y hoy sabemos que los eucariotas —es decir, nosotros— surgimos de una simbiosis entre una arquea y una bacteria.
3. Eucariotas: la rama compleja
- Origen: hace unos 2.000 millones de años, una arquea hospedadora incorporó una bacteria aerobia que se transformó en la mitocondria. Este evento de endosimbiosis fue el salto cualitativo que dio lugar a las células eucariotas.
- Características: poseen núcleo, organelos internos y un citoesqueleto complejo.
- Impacto: de ellas derivan todos los organismos pluricelulares —plantas, animales, hongos— y, finalmente, la conciencia humana.
Un árbol enraizado en LUCA
La diversificación de LUCA nos muestra que la vida no es un progreso lineal hacia mayor complejidad, sino una red divergente. La mayoría de la diversidad biológica sigue estando en bacterias y arqueas; los eucariotas son apenas una rama tardía, pero la que dio origen a la complejidad multicelular.
LUCA, entonces, no es solo un ancestro: es la raíz del árbol de la vida. Desde esa raíz, tres grandes ramas se abrieron paso, colonizando todos los rincones del planeta y dejando claro que la evolución es un proceso de exploración múltiple, no un destino único.
La vida en el Cosmos
El estudio de LUCA y de la evolución química en la Tierra primitiva no es solo una mirada arqueológica al pasado, sino también una guía para el futuro de la ciencia. Si logramos reconstruir cómo surgió la vida aquí, podemos definir qué señales buscar y dónde buscarlas en otros mundos.
1. Condiciones universales
Los escenarios que dieron origen a la vida en la Tierra —agua líquida, moléculas orgánicas, fuentes de energía química— no son exclusivos de nuestro planeta. Se han detectado aminoácidos en meteoritos como el Murchison, y moléculas orgánicas complejas en las nubes interestelares. Esto sugiere que la química prebiótica es universal y puede repetirse en otros lugares del cosmos.
2. Planetas y lunas candidatos
- Marte: evidencia de agua pasada y presencia de moléculas orgánicas detectadas por la misión Curiosity.
- Europa (luna de Júpiter): océano subterráneo bajo una corteza de hielo, calentado por la marea gravitacional.
- Encélado (luna de Saturno): géiseres que expulsan agua con compuestos orgánicos desde un océano interno.
- Exoplanetas: telescopios como James Webb comienzan a detectar atmósferas en planetas de otras estrellas, buscando trazas de agua, metano u oxígeno.
3. Biofirmas y límites de la vida
- Al estudiar los metabolismos de bacterias y arqueas, heredados de LUCA, identificamos las biofirmas mínimas: gases como metano, óxidos de azufre o huellas de actividad metabólica.
- Además, los extremófilos modernos —arqueas que viven en volcanes, bacterias que prosperan en hielo o radiación— amplían nuestra noción de dónde puede existir vida, más allá de la “zona habitable” clásica.
Marte: un eco de la Tierra primitiva
Esta semana, la NASA anunció uno de los hallazgos más sólidos hasta ahora: el rover Perseverance encontró en el cráter Jezero rocas con moléculas orgánicas, minerales asociados a procesos biológicos (fosfatos y sulfuros de hierro) y patrones químicos que podrían ser biofirmas. Los investigadores lo calificaron como la señal más clara hasta la fecha de que Marte pudo haber albergado vida microbiana hace miles de millones de años.
Lo revelador es la similitud con lo que sabemos de la Tierra primitiva. Jezero fue un lago antiguo, alimentado por ríos, un ambiente de agua líquida y minerales: exactamente el tipo de escenario donde aquí se forjó la evolución química que culminó en el RNA. Si en la Tierra la vida surgió rápido, antes incluso del Bombardeo Intenso Tardío, no es descabellado pensar que Marte, en el mismo periodo, haya transitado caminos parecidos.
Por ahora, no hay pruebas definitivas de vida marciana, solo indicios poderosos. Pero incluso si esos patrones resultan ser puramente geológicos, nos recuerdan que los ingredientes y ambientes necesarios para la biogénesis no son exclusivos de la Tierra. La pregunta que plantean estos hallazgos no es ya si Marte tuvo agua, sino si también tuvo su propio “LUCA” —un ancestro perdido que nunca llegó a prosperar— o si tan solo fue un ensayo cósmico fallido de la vida.
Reflexión filosófica: la universalidad de la vida y el silencio del cosmos
Los descubrimientos recientes en Marte y las reconstrucciones de la Tierra primitiva apuntan a una conclusión audaz: la vida no es una rareza cósmica, sino una consecuencia probable de la química universal. Aminoácidos en meteoritos, moléculas orgánicas en nubes interestelares, agua en lunas heladas: todo sugiere que el universo está sembrado de los ingredientes y procesos que, aquí, condujeron a LUCA. Si en la Tierra la vida surgió en apenas unos cientos de millones de años, ¿por qué no habría de repetirse en otros mundos?
Pero aquí aparece la paradoja de Fermi: si la vida es común, y si algunas de esas formas habrían tenido miles de millones de años para evolucionar hacia la inteligencia tecnológica, ¿por qué el cosmos nos devuelve un silencio tan absoluto? ¿Dónde están todos? decía Fermi.
La tensión entre ambas visiones es desconcertante. Por un lado, todo indica que la vida simple es un fenómeno casi inevitable en condiciones adecuadas; por otro, la ausencia de señales de civilizaciones avanzadas nos recuerda que la vida compleja e inteligente puede ser extraordinariamente frágil o improbable. Tal vez la vida surja con frecuencia, pero quede atrapada en formas microbianas, incapaz de dar el salto hacia sociedades tecnológicas. O quizá civilizaciones enteras hayan florecido y desaparecido antes de poder dejar huellas duraderas.
Filosóficamente, esta contradicción nos obliga a reconocer la diferencia entre la universalidad de la biología y la rareza de la conciencia. El universo puede estar lleno de microbios, algas y ecosistemas simples, mientras que la emergencia de seres capaces de preguntarse por su origen podría ser un acontecimiento casi único. LUCA sería común en muchos planetas; Sócrates, Darwin o nosotros, quizá no.
Así, el hallazgo de huellas de vida en Marte no resolvería la paradoja de Fermi, pero sí la profundizaría: confirmar que la vida es universal haría aún más misterioso el silencio de las estrellas.
Colofón
El viaje que va de Aristóteles a LUCA, de la generación espontánea a los meteoritos carbonáceos, del mundo de RNA al DNA y de allí a la reconstrucción de nuestro ancestro común, muestra una constante: la vida es un fenómeno que no necesita dioses ni milagros, sino tiempo, química y condiciones propicias.
Hoy sabemos que la vida surgió en la Tierra en un marco de cataclismos, que sobrevivió al Bombardeo Intenso Tardío y que dejó en LUCA la raíz de todo lo que respiramos, pensamos y somos. Sabemos también que los ingredientes de la biología se encuentran en el espacio, que Marte conserva huellas tentadoras y que otros mundos guardan océanos ocultos bajo su hielo. La conclusión parece inevitable: la biología es universal.
Y, sin embargo, el silencio del cosmos nos recuerda que la conciencia es rara. Tal vez la vida simple sea la norma del universo, pero la vida que reflexiona sobre sí misma —la que escribe, recuerda y se pregunta por su origen— sea la excepción. En esa tensión se cifra nuestro destino: somos, a la vez, hijos de un proceso común a toda la materia y herederos de una improbable chispa de conciencia.
Mirar hacia atrás, hacia LUCA, es descubrir nuestra raíz. Mirar hacia adelante, hacia las estrellas, es reconocer que la vida quizá abunde, pero que la conciencia, tan frágil y preciosa, podría ser el verdadero milagro. Y si eso es cierto, nuestra responsabilidad es doble: preservar la vida y honrar la conciencia, porque en ellas se cifra no solo nuestra historia, sino también nuestra unicidad en el universo. FIN


